将合成 β-聚羟基丁酸酯 (PHB) 的酶基因,λ-噬菌体裂解细胞的酶基因及能合成血红蛋白VHb 的酶基因——合成透明颤菌血红蛋白酶基因Vgb,共同转入 E. coli 中,发酵产聚 β-羟基丁酯(可降解的高分子) 工程菌 PHB 产量达 194 g/L,占细胞干重高达 90%,同时解决了破
壁及高密度培养的生化工程问题,已进行投产[5]。
抗体酶,又叫催化抗体,具有抗体高选择性和酶的高效催化性。谷胱甘肽过氧化物酶(GPX) 可以清除体内活性氧 (ROS) 所带来的细胞损伤,治疗炎症、糖尿病和心血管病等。根据 Pauling 过渡态理论,设计了多种含硒的谷胱甘肽过氧化物酶模拟物,通过杂交瘤技术免疫小鼠,化学诱变成功制备了分子量小能穿透细胞膜有治疗价值的具 GPX 活力的单链抗体模拟物,为天然兔肝酶活力的 8.5 倍。用人源基因替换鼠非保守区,全基因片段在噬菌体展示,构建了人源化 GPX 单链抗体酶,还制备了含硒多肽 GPX 模拟物,活力为 68.7 U/mg,为天然酶的 80%,最适 pH 和温度与天然酶相近。通过鼠表皮细胞抗氧化实验,具强抗氧化能力。用化学修饰将 θ 型谷胱甘肽硫转移酶 (GST) 转化为含硒 GPX 酶,能有效还原 H2O2,催化能力为国外临床试验的PZ51 (含硒杂环化合物) 的5 000倍[5]。
通过分子印迹技术,变性卵清蛋白作为印迹分子,除去印迹分子后,印迹分子结合部位中 Ser被 Sec 取代,该蛋白分子有高的 GPX 活力,为PZ51 的 820 倍[5]。还合成一系列环糊精衍生物的有机硒化合物作为 GPX 模拟物,对膜通透性好,无免疫原,体内半衰期长[5]
。GPX 模拟物6-ImTeCD 催化谷胱甘肽还原 H2O2活力为6.8 U/μmol,还原 t-BuOOH 级 CuOOH 活力是 9.7和 15.2 U/μmol
[10]。
核酶 (Ribozyme) 是小分子 RNA,也是一种多功能催化剂,可催化病毒 RNA 自我切割或断裂反应。我国亦进行了[R-]锤头核酶基因防治马铃薯纺锤病毒 (PSTVd) 和[R]锤头核酶基因防治椰菜花叶病毒 (CaMy) 的工作
[1]
。
此外,还有利用基因工程菌丝氨酸羧甲基转移酶生产 L-丝氨酸[4]
。重组毕赤酵母生产腺苷甲硫氨酸药物[5]
。重组菌生产 1,3-丙二醇及2,3-丁二醇[8]
。构建 E. coli 工程菌生产色氨酸[8]
。
芽短梗霉工程菌生产聚苹果酸[10]
。芽胞杆菌突变株 Y89D 生产环糊精[8]
。重组 E. coli 合成抗禽流感药物达菲前体草莽酸[10]
。枯草芽胞杆菌突变株生产苯丙氨酸。利用菌株基因敲除提高工程菌生产化工产品原料丁二酸产量[10]
。大肠杆菌工程菌全细胞转化 L-苯丙氨酸生产 α-苯丙酮酸[10]。
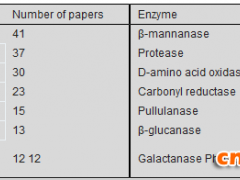
酶工程会议还有许多酶基因克隆与表达的论文,不能一一叙述,如:修复 DNA 损伤的O6-甲基鸟嘌呤 DNA 甲基转移酶,甜菜碱醛脱氢酶提高抗氧化酶活力,与噬菌体蛋白同源的新核酸酶。甜蛋白基因的表达,蓝藻蛋白裂合酶,抗虫胆固醇氧化酶,嘧啶核苷磷酸化酶[4],乙醇氧化酶,合成 D-氨基酸的 N-氨甲酰-D-氨基酸酰胺水解酶[5],辣根过氧化物酶[7],烟酸脱氢酶[8],乳糖酶[7],甲烷单加氧酶,苯乙烯单加氧酶,右旋糖酐蔗糖酶[8],脂肪醛去甲酰加氧酶,多聚乙酰合成酶,CO2关键酶与限速酶,耐热羧酸酯酶,硫氧化还蛋白谷胱甘肽还原酶[10],烟酸脱氢酶[7],毕赤酵母 SASA 冠状病毒的表达,癌转移有关的人基质金属蛋白酶,重组毕赤酵母表达人血清白蛋白,耐热单加氧酶,合成 r-亚麻酸的Δ6-脱氢酶在酵母中表达,E. coli 表达1,3-丙二醇氧化还原酶生产多聚纤维单体 1,3-丙二醇[5]
,壳聚糖酶,酰胺酶,甘油脱水酶,转谷氨酰酶,丙烯腈水解酶,嗜热菌角质酶-CBD 融合蛋白[11],人细胞色素 P450酶在 E. coli 中表达,耐碱性葡萄糖脱氢酶在 E. coli 中表达,丙氨酸消旋酶[8]
,果胶内切水解酶,手性药物合成的NADPH 酮基还原酶,酪氨酸激酶,糖基合成酶,天冬酰胺合成酶,S-腺苷同型半胱氨酸水解酶,丙酮酸脱氢酶,无机焦磷酸酯酶[6],E. coli 表达治疗肝癌与黑色素瘤的精氨酸脱亚胺酶。制备手性醇的酮还原酶。热稳定制备芳基醇的醇脱氢酶[9]。异源表达 β-1,3-氮乙酰胺基葡萄糖转移酶制备人乳寡糖,克隆了 5 种糖核苷酸合成的关键酶合成相应的糖核苷酸,E. coli 表达细菌脂肪氧合酶用于绿色香料和植物激素合成。E. coli诱导表达对治疗癌症及高半胱氨酸症有价值的深海甲硫氨酸 r-裂解酶。E. coli 表达 Bst DNA聚合酶[9]。抗生素骨架聚酮合成的迭代聚酮还原酶。肿瘤标记物环氧化酶-2 的人源单链抗体。
E. coli 表达双功能 D-乳酸脱氢酶,重组表达系列琼胶酶制备琼胶寡糖。里氏木霉异源表达木质素过氧化物酶[10],构建了 α-乙酰乳酸脱羧酶啤酒酵母工程菌用于啤酒生产[4]。聚酮合酶是最强大的化学合成酶[10],正在进行结构功能研究。
2 酶与微生物细胞的生物合成与催化传统的化学催化已不能满足可持续发展的需要,高立体选择性的微生物细胞或酶已成为手性生物合成的亮点。在这 25 年中,我国工业生物催化已成为具有相当规模和技术水平的产业,在世界上占有举足轻重的地位。
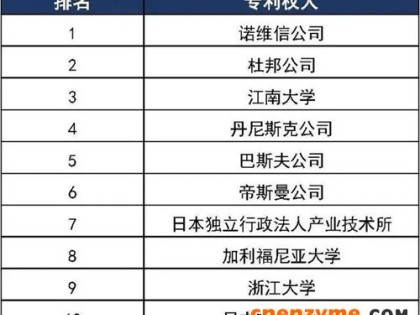
在第一至九届全国酶工程会议的 978 篇论文中,基因工程酶方面的有 356 篇,生物催化合成论文有 186 篇,占第二位。其中脂肪酶催化合成论文占 55 篇,占生物合成的第一位。
我国在有机相、离子液体、非水溶剂系统已成功利用脂肪酶进行了手性 2-辛醇[3,5,7-8]拆分,(R,S) 仲辛醇拆分[3,5-6],α-苯乙醇拆分[6],酮洛芬乙烯水解拆分[10],合成己酸乙酯[3-4,7,11],合成辛酸乙酯[7],油酸乙酯[7],庚酸乙酯[10],癸酸乙酯[7],生物柴油[7-9,11],脂肪酸甲酯[11],二元酸酯[8],单甘油酯[4],油酸油醇酯[3],香叶醇酯[3],油酸酯[3],乙酸乙烯酯[3],脂肪酸糖脂[9],(S)-α-氨基-3-苯氧基苄醇[5],1,3-丙二醇羧酸酯[3],棕榈异辛酯[8],癸酸甘油酯[5],维生素 A 棕榈酸酯[5],抗坏血酸脂肪酸酯[5],蔗糖酯[7],聚羟基丙酸酯[7],葡萄糖月桂酸单酯[8,11],果糖月桂酸单酯[8],及 (+)反式菊酸[4],以及新闻纸脱墨[7,10]等。
细胞展示技术是将外源蛋白锚定在细胞壁上,酶分子的固定化可提高酶的稳定性和对有机溶剂的耐受性,毕赤酵母对外源脂肪酶表面展示系统可催化合成果糖脂、己酸乙酯,葡萄糖酯、生物柴油、月桂酸单糖酯、脂肪酸甲酯[8,11]等。离子液体体系是新型绿色非水溶剂,酶法微藻藻油生产生物柴油达 90%转化率。并合成了
脂肪酸糖脂[9-10]。
利用酯酶催化合成酒香己酸乙酯[3],单一性2-辛醇拆分合成抗癌药物前体 (2S,3R)-3-苯基缩水甘油酸甲酯[10],固定化合成脂肪族聚酯
[10],
菌株不对称合成 L-薄荷醇[7],拆分 1,1,1-三氟-2-辛醇[10],有机合成壬酸香草醇酯 (辣椒素脂)[8]
。
用蛋白酶与化学法相结合,在低水有机相中合成了 4 种含 Arg-Gly-Asp(RGD) 的细胞粘附 肽 RGDS 、 RGD(NH2)2、 RGE-NH2和RGDS-NH2,有抗肿瘤作用[7]
。
目前,已知酶有 4 428 种 (至 2011 年),工业上用酶大多数为水解酶,但氧化还原酶在酶中比例却约占 32%,很多氧化还原酶未能被开发
,因此拓展氧化还原酶是工业技术中一项重要内容,但需解决辅酶循环再生的难题。为建立微生物细胞氧化还原酶催化体系,提高内源辅酶
利用率,我国已利用 E. coli 重组菌双辅酶底物偶联固定化面包酵母羰基还原酶合成 (S)-苯基乙二醇手性模块物[8]
。用双羰基还原酶偶联辅酶再生体系合成他汀类药物手性侧链——3R,5S二羟基-6-苄基己酸乙酯[8]
。重组菌羰基还原酶合成医药合成用的手性醇 (S)-4-氯-3-羟基丁酸乙酯,(R)-邻氯扁桃酸甲酯和 (S)-2-羟基-4-苯基丁酸乙酯[9]。用烯酮/烯酯还原酶还原 C=C 双键合成有商品价值的底物,对甲基马来酰亚胺、R-香芹酮、衣康酸二甲酯、乙烯酰氧基丙烯酸甲酯、茶香酮、苄烯丙二腈都有较高的转化率[10]
。用辅酶再生体系酵母醇脱氢酶不对称合成医药,农药光学手性醇中间体 (R)-扁桃体酸甲酯和 (R),(S)2-辛醇[8]
。菌株 D-氨基酸脱氢酶和胺氧化酶合成手性胺 (如 D-丙氨酸、D-苯丙氨酸、D-叔亮氨酸)。已建立了 D-氨基酸合成的酶催化平台[10]
。氧化葡萄糖酸杆菌高选择性氧化合成工业上第二位的芳香醛——苯甲醛[8]
。嗜热酮还原酶辅酶再生系统不对称合成医药中药中间体 S-1-苯基 222 三氟乙醇[9]
。E. coli 共表达全细胞甲酸脱氢酶催化茴香硫醚不对称合成手性亚砜——甲基苯基亚砜[9]
。金黄杆菌烯醇还原酶催化 C=C 双键不对称还原环状烯酮产生光学纯烷烃化合物用于手性中间体合成[10]
。金黄杆菌短链脱氢酶不对称合成匹瑞匹坦药物中间体(R)-[3,5-二 (三氟甲基) 苯基]乙醇[10]
。芽胞乳杆菌 D-乳酸脱氢酶催化谷氨酸合成 α-酮戊二酸[10]
。高活性全细胞醇脱氢酶不添加辅酶合成米格列醇[6]
,基因敲除提高酶表达量构建甘油脱氢酶合成 1,3-二羟基丙酮 (最简单的多羟基酮糖)[8]
。利用天然多脱氢酶体系甲烷氧化细菌生物催化二氧化碳制甲醇[5]
。厦门大学挖掘了海洋微生物的8 种氧化还原酶体系[9]
。
国内已形成生物催化与微生物转化的科研开发群体,江南大学与浙江鑫富生化公司合作,用产 D-泛解酸内酯水解酶的串珠镰孢菌水解拆分
得到光学纯的 D-泛解酸内酯,成功地用于 D-泛酸钙和 D-泛醇的生产,D-泛酸钙产量全球第一[5,10]
。此外,还固定化诺卡氏菌水解顺式环氧琥珀酸生产 L-酒石酸[7]
。微生物转化生产香草酸与香草醛,又名香兰素 (3-甲氧基-4-羟基苯甲醛) 高档香料[5]
。转化巴豆甜菜碱制 L-肉碱[4]
,
采用双水相合成用于倍他司汀抗组胺药物手性中间体-(S)-(4-氯苯基)-(吡喃-2-基) 甲醇[9]
。定向进化E. coli菌株精氨酸脱亚胺酶为治肝癌药物[9]
。
融合酶是利用融合蛋白将几种功能蛋白集成于一体的嵌合体。构建肝素酶的双重融合蛋白体系,如麦芽糖结合蛋白融合或荧光蛋白融合,促
进了肝素酶的可溶性表达,用于生产低分子量抗栓药物——肝素[6-7,9,12]
。双亲短肽融合
环糊精糖基转移酶的 N 末端构建 6 种融合酶合成克服 Vc 氧化的 2-氧-D-吡喃型葡萄糖基-L-抗坏血酸[10]
。固定化自组装短肽 18A 与腈水解酶融合表达生产扁桃腈[10]
。用嗜热子囊菌角质酶-CBD 融合蛋白精炼棉织物[8]
。